论文推荐 | 甘薯淀粉合成与降解糖化的研究进展

甘薯淀粉合成与降解糖化的研究进展
覃晓晖1,李堯鑫1,方浩宇1,余鹏波1,黄鸿文1,
陈选阳1,2,3,廖素凤1,2,3,*杨志坚1,2,3

作者单位
1. 福建农林大学 农学院
2. 福建农林大学 海峡两岸农业技术合作中心
3. 福建农林大学 作物遗传育种与综合利用教育部重点实验室
引用格式
覃晓晖,李堯鑫,方浩宇,等.甘薯淀粉合成与降解糖化的研究进展[J].农产品加工,2026,(10):94-102+109.
基金项目
福建省科技厅产学合作项目(2025N5003);广州市重点研发计划项目(2025B03J0007)



一、引言
甘薯(Ipomoea batatas L.)是世界范围内重要的粮食作物,具有高产、适应性强、用途广泛等特点,是保障国家粮食安全与功能性食品的重要资源。甘薯富含碳水化合物、膳食纤维、多种维生素、矿物质及多种对人体有益的成分,适量食用甘薯可提高人体免疫力,保持身体健康。得益于甘薯丰富的营养成分和多种健康益处,世界卫生组织(WHO)曾将甘薯列为最佳食物。
甘薯块根的加工性能和食用品质受多种因素的综合影响,其中直链淀粉与支链淀粉的比例、淀粉粒径及结晶结构是决定甘薯淀粉结构特性的关键因素,并影响淀粉的糊化特性、水溶性及膨胀力。淀粉和可溶性糖含量也直接影响甘薯的食味品质与加工特性。淀粉是甘薯块根中的主要成分,而可溶性糖又主要来源于淀粉的降解,在甘薯的生长阶段,淀粉作为主要的能量储备物质需要被大量合成,而在采收后,块根需将淀粉降解为可溶性糖,以满足鲜食或加工对糖度的需求。然而,采收后的甘薯块根中,淀粉仍然占据主导地位,淀粉向可溶性糖的转化效率相对较低,导致可溶性糖含量不足,进而影响其风味和口感。尤其是在夏秋季节采收的甘薯,由于收获时温度较高,块根内的淀粉合成代谢活动仍然活跃,导致甘薯块根中淀粉含量较高,而可溶性糖含量较低,难以满足直接食用所需的甜度和食品加工的糖含量标准。为了提高甘薯的食用和加工价值,对其进行糖化处理,即将块根中大量的淀粉转化为可溶性糖等甜味物质,是一种有效的手段。传统依赖自然贮藏的糖化方式因周期长、损耗率高、糖分积累不可控等原因,严重制约了甘薯产业链的时效性与经济性。
深入研究甘薯快速糖化条件,对于提升甘薯加工的水平具有重要意义。目前,关于淀粉代谢的研究主要集中在淀粉生物合成的分子机制和遗传基础等方面。但对于淀粉降解过程中,淀粉转化为可溶性糖类物质的潜在调控机制,仍有待深入研究。通过阐明甘薯淀粉降解糖化的生理机制及分子调控靶点,为实现采后甘薯高效糖化与优质品种选育提供理论基础。

二、甘薯淀粉特性
(一)直/支链淀粉含量及其对甘薯食味品质的影响
淀粉是甘薯块根的主要成分,占干质量的50%~ 80%。淀粉的存在形式为葡萄糖聚合体,是D -葡萄糖通过α-1,4和α-1,6糖苷键连接而成的高聚物,根据葡萄糖分子间的连接方式不同分为直链淀粉和支链淀粉。直链淀粉是由α-1,4糖苷键连接线状大分子,而支链淀粉包括主链和支链,主链由α-1,4糖苷键连接,支链由α-1,6糖苷键连接。甘薯块根中总糖、直链淀粉和支链淀粉质量分数分别为1.8%~ 4.7%,15.3%~31.2%和68.8%~84.7%淀粉中直链淀粉和支链淀粉的比例对甘薯食用品质、加工特性等有着直接影响支链淀粉具有较好的黏着性能,支链淀粉含量高的甘薯口感偏软糯性;而直链淀粉的黏着性差,不易形成黏稠的质地,其含量高时甘薯口感偏粉。
(二)淀粉颗粒对甘薯加工品质的影响
淀粉颗粒的粒径大小与其膨胀势、黏度和流变性等特性有关,直接影响甘薯加工品质。支链淀粉具有高度分支结构,其分支特性有利于淀粉颗粒的膨胀和有序堆积,通常与较大的颗粒尺寸相关;而直链淀粉则通过氢键形成致密结构,可能限制颗粒膨胀,导致粒径较小。因此,直链淀粉含量越高,淀粉颗粒的粒径越小;支链淀粉含量越高,则淀粉颗粒的粒径越大。淀粉颗粒的粒径大小与淀粉糊化程度紧密相关,即粒径越小,淀粉糊化程度越高,糊化速率也越快,同时,小颗粒淀粉因比表面积更大,更易被酶解。直链淀粉含量与糊化焓呈显著正相关,即直链淀粉含量越高,淀粉粒径越小,糊化焓越高。淀粉颗粒的大小也会直接影响甘薯淀粉面条的加工性能和成品质量,由小粒径颗粒(<20 μm)制成的干淀粉面条和熟淀粉面条的加工性能远优于由大粒径颗粒淀粉制成的面条。研究表明,支链淀粉中的长链更易形成稳定的双螺旋结构,需要更高的能量才能破坏,故长链比例越高,甘薯淀粉糊化温度越高;反之,支链淀粉的短链比例较高时,糊化温度则较低。
(三)淀粉改性对甘薯加工品质的影响
甘薯淀粉的某些天然特性限制了其直接应用,因此对其进行改性可提升加工品质、拓宽应用范围。对甘薯淀粉的改性主要是通过物理、化学、酶法和复合方法进行。物理改性主要是通过温度、脉冲电场、挤压和辐射等方法;化学改性是指通过化学试剂与淀粉分子中的羟基发生反应,从而改变淀粉的物理化学性质,包括醚化、乙酰化、酸改性、阳离子连接、酯化、交联和氧化;酶法改性主要是通过淀粉水解酶和淀粉分支酶对淀粉的结构,特别是支/直链进行修饰。
温度处理包括烘烤、超高温加热、热湿、退火处理等,可改变淀粉的结晶结构和糊化特性、提高淀粉的抗老化性。利用高压电脉冲(PEF)技术可诱导淀粉颗粒的结构和化学变化,从而改善糊化、溶解度、黏度和溶胀能力等特性,改性后的淀粉可更好地保留营养和感官品质,同时还增强其加工性能。电子束处理能够影响甘薯淀粉的结构、理化性质和体外消化特性,随着电子束辐照剂量的增加,相对结晶度、分子量、长链比例、糊化熵、黏度和溶胀力降低,短链比例、溶解度和抗性淀粉含量提高。利用紫外线、微波、伽马射线等微波辐射可使淀粉受热均匀,提高改性效率。将烷基等基团引入淀粉分子中可实现淀粉的醚化,淀粉的羟丙基化反应可提高淀粉的溶解性和糊化稳定性。通过辛烯基琥珀酸酐(OSA)可对甘薯淀粉进行酯化改性,使甘薯淀粉具有更高的黏度、更好的糊透明度和更低的逆老化程度。利用乙酸酐、己二酸酐等对甘薯淀粉进行酯化改性,可降低淀粉的糊化温度,提高淀粉的冻融稳定性。二羧酸等交联剂能在淀粉分子间形成化学键,提高淀粉的耐热性、耐酸性和耐剪切性,交联淀粉在食品工业中常用作增稠剂和稳定剂。使用次氯酸钠等氧化剂氧化淀粉,可降低淀粉的黏度,提高其成膜性,氧化淀粉常用于食品添加剂,改善食品的质地和稳定性。糖基转移酶将糖基转移到淀粉分子上,改变淀粉的结构和性质;使用淀粉蔗糖酶和支链淀粉酶处理淀粉,可产生具有更长线性链的淀粉,有利于与不饱和脂肪酸形成复合物,从而控制淀粉的消化性,并提高其抗氧化稳定性。

三、甘薯淀粉合成
(一)淀粉合成关键酶
叶绿体和淀粉体是植物淀粉合成的场所,甘薯块根中淀粉合成的主要场所是淀粉体。叶片中的叶绿体负责贮藏光合作用中合成的淀粉,而淀粉体则是贮藏淀粉合成的场所。在甘薯块根中,光合作用产生的蔗糖通过韧皮部运输至淀粉体,经蔗糖转运体(SUT)进入后,通过糖酵解途径转化为葡萄糖- 1 -磷酸(G1P),最终在ADP -葡萄糖焦磷酸化酶(AGPase,EC 2.7.7.27)的催化下生成淀粉合成的直接前体——腺苷二磷酸葡萄糖(ADPG),ADPG作为葡萄糖供体,将葡萄糖残基转移到α-1,4-葡聚糖链的非还原端,延长淀粉分子。ADP -葡萄糖焦磷酸化酶(AGPase)催化ADPG生成的反应是淀粉合成的限速步骤。淀粉合成酶是淀粉合成的关键酶,负责催化α-1,4-葡聚糖链的延伸,主要包括颗粒结合型淀粉合成酶(GBSS,EC 2.4.1.242)和可溶性淀粉合成酶(SS,2.4.1.21)等多种亚型。GBSS催化直链淀粉的合成,GBSS以ADPG为底物,将葡萄糖基连续添加至淀粉链的非还原端,形成线性α-1,4糖苷键链,进而形成线性直链淀粉。SS参与支链淀粉的合成,并通过可溶性淀粉合成酶(SS)、淀粉分支酶(SBE,EC 2.4.1.18)和异淀粉酶(ISA,EC 3.2.1.68)的协同作用合成支链淀粉。SS又有多个同工酶,包括SSI,SSII,SSIII和SSⅣ,其中SSI优先延伸短链,形成支链淀粉的外侧短链;SSII延长中等链长,形成支链淀粉的B1链,即内侧中等链,与SSI协同作用,影响支链淀粉的簇状结构;SSIII调控支链淀粉的超长链合成,影响淀粉颗粒的整体结构;SSⅣ主要参与淀粉颗粒的起始和初期形成,可能调控淀粉合成的起始步骤。淀粉分支酶(SBE,EC 2.4.1.18)通过切割α-1,4键并重新连接α-1,6键引入分支点,形成支链淀粉的骨架。异淀粉酶(ISA,EC 3.2.1.68)负责修剪过度密集或非结晶区的α-1,6键,优化支链结构,确保淀粉颗粒的结晶完整性。
淀粉合成网络见图1。
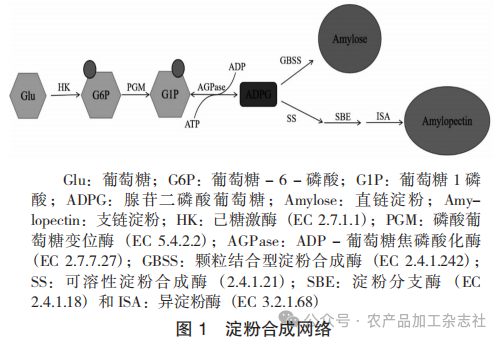
(二)淀粉合成的分子调控网络
甘薯淀粉合成受多层级调控网络控制,涵盖前体代谢、限速酶活性、碳源运输及转录因子调控,各模块协同决定淀粉产量与品质。
1. 前体代谢与淀粉合成
甘薯淀粉前体代谢酶与修饰蛋白的协同作用精细调控合成。在甘薯贮藏根中,己糖激酶(HK,EC 2.7.1.1)催化葡萄糖生成葡萄糖- 6 -磷酸(G6P),随后由质体型磷酸葡萄糖变位酶(IbPGM,EC 5.4.2.2)介导G6P与葡萄糖- 1 -磷酸(G1P)的可逆转化。有研究表明,在低淀粉甘薯品种中IbPGM的过表达可显著提高贮藏根中淀粉含量,同时降低蔗糖水平,表明IbPGM可能通过增强G1P供应正向调控淀粉积累。甘薯谷胱甘肽S -转移酶IbGSTL2(EC 2.5.1.18)被发现与IbPGM发生蛋白互作,其过表达不仅使支链淀粉短链比例增加、小型晶体比例上升、结晶度提升,更通过提高分支密度使糊化温度降低。
2. AGPase结构与淀粉合成
腺苷二磷酸葡萄糖焦磷酸化酶(AGPase,EC 2.7.7.27)作为淀粉合成的关键限速酶,催化G1P与ATP生成ADPG。甘薯AGPase被发现是一种异源四聚体,其中小亚基IbAPS被证实是酶活性的主要调控者,过表达IbAPS可提高AGPase活性1倍,使块根淀粉含量显著增加,同时通过提高光合效率,增加淀粉合成相关酶活性来提高甘薯产量。进一步研究发现,CBF/DREB家族转录因子StTINY3通过双重机制调控淀粉-蔗糖平衡,一方面直接激活AGPase大亚基基因StAGPaseL3和葡萄糖- 6 -磷酸转运蛋白基因StG6PT2的表达,促进淀粉前体合成;另一方面,上调转化酶抑制因子StInvInh2,抑制蔗糖水解酶活性,从而将碳流定向导入淀粉合成途径。蔗糖非发酵相关蛋白激酶IbSnRK1通过磷酸化修饰AGPase和可溶性淀粉合酶(SSS),可提升淀粉含量,下降直链淀粉比例,并提高淀粉颗粒尺寸分布集中度和结晶度。
3. 跨膜转运与质子梯度驱动的碳流优化
蔗糖从源叶到贮藏根的高效运输依赖于跨膜质子梯度的建立与维持。液泡膜H+-焦磷酸酶(IbVP1,EC 3.6.1.1)位于液泡膜,通过水解清除液泡内的焦磷酸(PPi),解除其对淀粉合成限速酶AGPase的抑制,同时激活AGPase、GBSSI等基因表达,直接促进淀粉合成。与之协同的质膜H+-ATP酶IbPMA1(EC 3.6.3.14)则通过建立胞外质子梯度,驱动蔗糖/ H+共转运体活性,加速蔗糖从源向块根库的运输,为淀粉合成提供充足碳源,其表达强度与块根淀粉积累速率呈显著正相关。二者在细胞内位置不同,形成空间分工,并共同作用,既保障了碳源的高效运输,又促进了碳源的高效转化,形成碳流定向运输的完整链条。
4. 转录因子调控淀粉合成
转录因子在调控甘薯淀粉生物合成中扮演着关键角色。IbNAC22被发现在高淀粉型甘薯品种普遍高表达,IbNAC22通过直接激活直链淀粉生物合成关键基因IbGBSSI的表达,同时抑制支链淀粉生物合成关键基因IbSBEI的表达,提高了淀粉和直链淀粉含量及淀粉凝胶化温度,但减少了颗粒大小、结晶度和支链淀粉链长分布。碱性亮氨酸拉链IbbZIP22在高淀粉含量的甘薯品种中具有更高的表达量,可能参与了淀粉的合成和代谢。Dof锌指转录因子SRF1被发现是通过对液泡转化酶基因的负调控,来调节贮藏根中的碳水化合物代谢。过表达SRF1会抑制液泡转化酶同工型的Ibβfruct2的表达,导致贮藏根的淀粉含量显著提高,而葡萄糖和果糖含量则大幅下降。

四、甘薯淀粉降解糖化
(一)淀粉降解
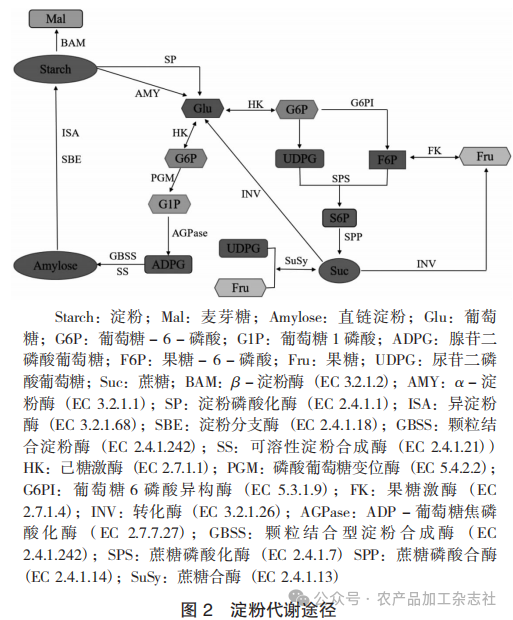
1. 淀粉降解的酶调控网络
甘薯块根中淀粉的降解方式包括水解和磷酸化降解。水解主要由α-淀粉酶(AMY,EC.3.2.1.1)和β-淀粉酶(BAM, EC 3.2.1.2)协同完成。磷酸化降解主要靠淀粉磷酸化酶(SP, EC 2.4.1.1)催化淀粉与无机磷酸之间的可逆反应,与水解降解不同,它不消耗ATP。AMY是一种内切淀粉酶,随机切割淀粉分子内部的α- 1,4 -糖苷键,生成低聚糖和麦芽糖;BAM作为一种外切淀粉酶,从非还原端依次水解麦芽糖单位,生成β-麦芽糖。值得注意的是,AMY对支链淀粉的分支点附近区域的α- 1,4 -糖苷键表现出更高的水解效率,而BAM则优先作用于直链淀粉的非还原末端[51-52]。SP通过磷酸化作用将淀粉从非还原端降解单个葡萄糖残基,并与Pi结合降解为葡萄糖- 1 -磷酸(G1P),基于定位差异,淀粉磷酸化酶又分为质体淀粉磷酸化酶(PHO1)和胞质淀粉磷酸化酶(PHO2)。
2. 蔗糖代谢调控网络
淀粉降解产物通过蔗糖合成酶(SuSy,EC 2.4.1.13)、蔗糖磷酸化酶(SPS,EC 2.4.1.14)和液泡转化酶(VIN,EC 3.2.1.26)进一步代谢。SuSy催化蔗糖与UDP -葡萄糖和果糖的可逆反应,其方向由底物浓度决定,在高蔗糖条件下,SuSy倾向于分解蔗糖生成UDP -葡萄糖和果糖;而在低蔗糖条件下,SuSy则催化UDP -葡萄糖与果糖合成蔗糖。SPS作为蔗糖合成的限速酶,催化UDP -葡萄糖与果糖- 6 -磷酸生成蔗糖- 6 -磷酸,后者经蔗糖- 6 -磷酸酯酶(SPP)水解生成蔗糖[55]。VIN则不可逆地将蔗糖分解为葡萄糖和果糖,其活性受液泡pH值和转化酶抑制剂(如INH1)的严格调控。
淀粉代谢途径见图2。
(二)低温糖化
低温处理是抑制甘薯发芽与减重、延长甘薯贮藏期、促进甘薯快速糖化的重要手段。低温处理激活了块根中淀粉降解途径,从而提高可溶性糖含量,提升鲜食口感并减少加工过程中额外的糖需求。当前,研究主要集中于低温贮藏对甘薯保鲜效果的影响和贮藏过程中抗氧化酶活性变化等方面。然而,针对低温诱导下甘薯淀粉降解与糖化的生理机制和分子机制的研究相对不足。
低温通过抑制淀粉合成与激活降解通路驱动甘薯糖化。在4 ℃低温条件下,淀粉合成途径的关键酶,如AGPase和GBSS的表达被抑制,而淀粉降解相关酶,如BAM、SPS和SS活性显著上调,导致淀粉大量转化为可溶性糖,增加细胞渗透势,以应对低温胁迫。研究表明,植物的耐低温能力与蔗糖含量呈显著正相关,拟南芥中过表达蔗糖合成酶基因AtSPS可显著提升耐寒性。低温贮藏条件下,耐冷甘薯品种块根中以酸性转化酶(AI)催化生成还原糖积累,不耐冷品种的块根则是以蔗糖合成酶(SuSy)催化的非还原糖积累。在番茄,黄瓜,和柑橘中,同样发现蔗糖的积累量与耐冷性呈显著正相关。
为促进甘薯低温糖化并防止块根受到冷害,多种物理和化学处理策略已被开发。研究发现,短期的低温贮藏:5 ℃下贮藏12 d,可促进蔗糖和绿原酸积累,提升甘薯甜度和抗氧化能力。梯度降温或昼夜温差可有效激活低温响应基因,降低冷害风险。LI X等人将甘薯块根先在10 ℃低温调节5 d,再转到4 ℃低温胁迫下贮藏,发现梯度低温贮藏既可有效增加甜味,同时可有效降低4 ℃贮藏产生的低温损伤。YUAN J等人]通过在1 d内25 ℃下12 h,12 ℃下12 h交替贮藏温度,在8 d的贮藏时间里,提升了β-淀粉酶(BAM)、蔗糖磷酸合酶(SPS)和蔗糖合酶(SS)活性,实现了蔗糖和果糖含量的大幅提升。此外,在40 kHz下外源超声处理,10 min可通过抑制多酚氧化酶(PPO)和过氧化物酶(POD)活性,延缓冷藏过程中的酶促褐变,同时提升超氧化物歧化酶(SOD)与过氧化氢酶(CAT)活性,增强抗氧化能力。
(三)高温糖化
目前,对甘薯高温糖化的研究集中在烹饪糖化、加热过程中,高温提高了α -淀粉酶和β -淀粉酶活性参与甘薯块根中淀粉酶水解,产生可溶性糖。蔗糖在新鲜甘薯中占总糖的49.92%~92.43%,而在熟薯中,麦芽糖含量最高,因此甘薯烹饪后的增甜物质成分主要是来自β-淀粉酶催化淀粉降解产生的麦芽糖,熟薯中的麦芽糖含量超过生薯的10倍糖含量显著影响蒸煮、烘烤甘薯的口感和甜度。
甘薯淀粉糊化开始的温度为70~74 ℃,α- 淀粉酶和β-淀粉酶是催化淀粉分解成麦芽糖最重要的酶,β-淀粉酶的活性在40~90 ℃内,随着温度的升高而先升高后降低,在60 ℃达到最高值,但高麦芽含量品类的甘薯体内的β -淀粉酶稳定性高于其他品类,高麦芽含量品类甘薯的淀粉糊化发生在较低温度下。甘薯块根含有α-淀粉酶和β-淀粉酶,二者在热稳定性、作用方式及催化特性方面具有显著差异,前者含量较低但热稳定性好,能直接作用于淀粉分子内部的α- 1,4 -糖苷键,水解葡聚糖苷链,生成糊精和短链寡糖,最适温度在70~ 75 ℃;而后者含量高、热稳定较差,且只能作用于非还原端,水解糊化的淀粉,释放麦芽糖单位。二者通过协同作用提升高温下的催化效率,α-淀粉酶首先破坏淀粉颗粒的晶体结构并产生大量非还原末端,为β-淀粉酶创造作用位点,由β-淀粉酶进一步降解极限糊精。甘薯干物质、淀粉、淀粉酶含量和淀粉酶酶活是决定不同品种甘薯高温糖化差异的重要影响因素,淀粉糊化温度越低,则糖转化效率越高。
通过转录组学分析探析高温处理下的甘薯块根转录调控网络,65 ℃高温可诱导甘薯中MYB、WRKY、bHLH转录因子家族基因出现显著性差异表达。在其他植物中,WRKY家族被证实参与植物的糖代谢和信号转导;WRKY转录因子通过结合糖响应元件调控糖代谢基因α-淀粉酶、β-淀粉酶的表达,参与糖信号转导与代谢平衡,影响植物糖积累与分配。在猕猴桃中,发现MYB和bHLH转录因子通过光响应参与淀粉代谢。

五、技术突破与挑战
(一)转基因和基因编辑在甘薯中的应用
国内外通过转基因技术调控甘薯中淀粉合成相关基因,如 ADP - 葡萄糖焦磷酸化酶(AGPase)大、小亚基基因、可溶性淀粉合酶Ⅰ(IbSSⅠ)基因、淀粉合酶Ⅱ(IbSSⅡ)基因、淀粉分支酶基因、质体磷酸葡萄糖变位酶(pPGM)基因等,可改变淀粉的含量、组成、颗粒大小和结构,从而影响甘薯的品质。通过转基因过表达参与甘薯淀粉合成的相关基因,如谷胱甘肽S -转移酶 IbGSTL2、蔗糖非发酵- 1相关蛋白激酶- 1基因(IbSnRK1),可增加甘薯淀粉含量并改善淀粉组成,提升加工品质。利用CRISPR/Cas 9介导的基因编辑技术,如编辑淀粉合成中的关键酶基因,可用于改良淀粉特性,提升加工品质。
(二)基因编辑在甘薯中的挑战
尽管转基因和基因编辑技术显著促进了淀粉合成和优化了糖代谢调控网络,但由于甘薯是6倍体,基因组大且复杂,多基因协同调控机制尚未完全解析。利用农杆菌或基因枪法的CRISPR递送系统效率不足,需同时编辑多个等位基因,脱靶风险高。实验室编辑的株系在田间可能因环境互作丧失预期表型。基因编辑甘薯的商业化目前仍面临法律法规的限制,尚未进入人们的餐桌。然而,随着全球对基因编辑作物监管政策的逐步放宽,如美国、日本对CRISPR作物的非转基因认定,以及消费者对精准育种技术接受度的提升,基因编辑甘薯有望在未来进入市场,为淀粉品质改良与功能食品开发提供新选择。
六、结语
系统性地介绍了甘薯淀粉结构与特性对加工特性的影响、淀粉生物合成及低温和高温下淀粉转化的生理和分子机制,通过总结前人对甘薯淀粉合成及降解糖化的研究成果,串联构建了淀粉代谢调控网络,为研究甘薯淀粉合成和快速糖化提供理论基础。对于甘薯的糖化机理,研究发现甘薯在低温和高温下糖化的酶调控网络具有较大不同,低温条件下以葡萄糖、果糖和蔗糖积累为主,而高温条件下,则是以麦芽糖积累为主,故通过高温和低温相结合的新型贮藏手段,将淀粉充分转化为可溶性糖,对甘薯快速糖化具有重要的研究意义。
在甘薯贮藏中,利用低温提升甘薯块根可溶性糖含量已经被多项研究证实,低温条件下淀粉代谢相关酶基因表达量和酶活性提升,如α-淀粉酶、β -淀粉酶、蔗糖合酶、蔗糖磷酸化酶、酸性转化酶等,淀粉合成相关基因如AGPase被抑制。直链淀粉是由α- 1,4糖苷键串联构成的线性葡聚糖,α-淀粉酶、β-淀粉酶、淀粉磷酸化酶均可直接作用于直链淀粉,并切断直链淀粉中的α- 1,4 -糖苷键,生成低聚糖、葡萄糖和麦芽糖,低温条件下这些酶的活性显著提升,有助于甘薯块根的可溶性糖积累,促进低温糖化。同时,在甘薯中探索并利用苹果WRKY32-Bam5 类似通路激活β-淀粉酶表达,可加速淀粉降解。同时,在其他植物的研究中也发现一些转录因子,ERFI-2、CBF1/2、WRKY32、MYB44等参与了低温下淀粉-糖的代谢调控。
淀粉酶是淀粉降解为可溶性糖的主要酶,在马铃薯中发现SbAⅠ这一淀粉酶抑制基因,通过抑制块茎中α-淀粉酶和β-淀粉酶的活性,有效阻断低温下的淀粉-糖转化,若在甘薯中去除类似抑制因子,可能加速淀粉降解速率,实现快速糖化。此外,通过外源激素,如赤霉素处理可定向提高β-淀粉酶活性,直接促进淀粉水解与还原糖积累。液泡转化酶是液泡中蔗糖转化成还原糖的重要催化酶,其活性直接影响块根中甜味物质的积累。马铃薯中发现转化酶是促进淀粉降解的主要酶类,而ERF-Ⅶ转录因子StRAP2.3可激活转化酶抑制剂基因StInvInh2表达,使马铃薯块茎还原糖积累减少,甘薯中可能存在的同源转录因子,或可通过沉默或基因编辑技术抑制其表达,进而促进块根中可溶性糖积累。同时,通过在甘薯中提升液泡转化酶表达,或许可促进还原糖积累,增加块根甜味。
高温条件下,主要是β-淀粉酶催化淀粉转化为麦芽糖的过程,酶稳定性和淀粉糊化特性决定了高温糖化的效果。通过酶改良工程改造α-淀粉酶和β-淀粉酶的热稳定性,开发耐高温的高效糖化酶制剂是实现甘薯快速糖化的一大策略。相关研究集中在高温烹饪下淀粉糊化特性、淀粉及糖类物质含量变化和淀粉酶活性变化情况,但对于高温条件下的贮藏特性研究甚少。
对于甘薯育种,可利用CRISPR/Cas9系统编辑淀粉降解关键基因,如BAM、SPS等,或开发温度响应型启动子驱动糖化酶表达,定向制造抗冷害-高糖化的甘薯品种;也可通过编辑 IbGSTL2与 IbInvInh2,培育“高淀粉-高甜度”功能型甘薯,兼顾加工品质与鲜食需求;也可通过过表达CBF/DREB转录因子增强低温下蔗糖合成能力,结合液泡膜糖转运蛋白TST1/2 同源基因的过表达,实现糖分液泡区室化存储,延长货架期。但是,甘薯多倍体特性让转基因的成功率和基因编辑的效率低下。未来,研究应聚焦6倍体基因组的精准编辑技术,同步靶向 AGPase、GBSS 等多拷贝基因,开发多靶点CRISPR系统,从而突破转化效率低的技术瓶颈。
编辑:刘 嘉
审核:关 颖
本篇文章刊登于《农产品加工》2026年5期下